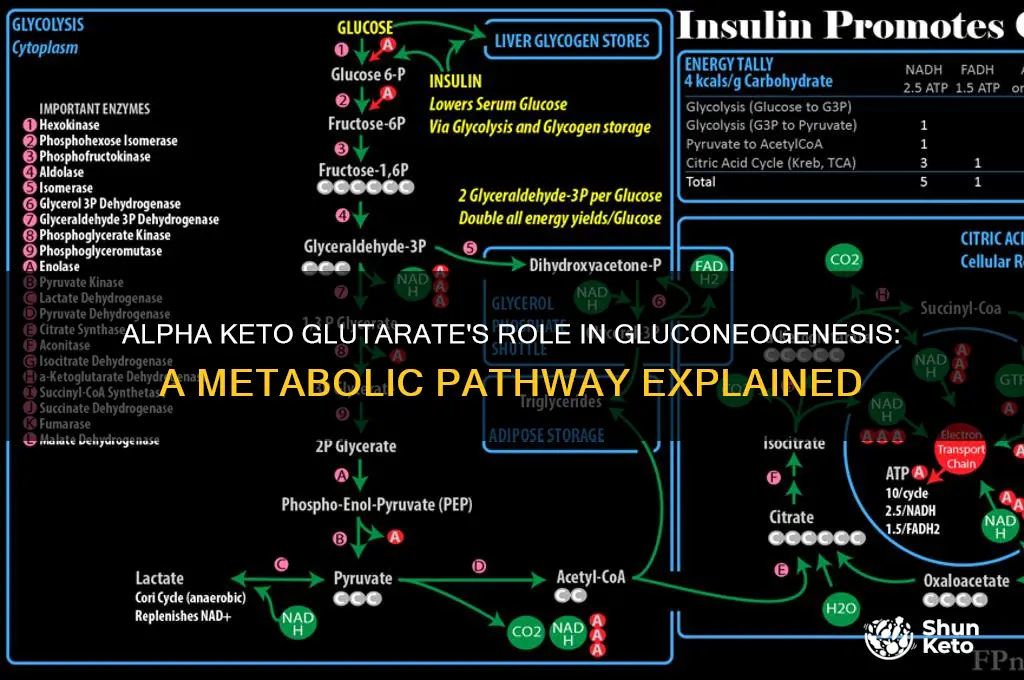
Alpha-ketoglutarate (AKG) plays a significant role in cellular metabolism, particularly in the context of gluconeogenesis, the process by which glucose is synthesized from non-carbohydrate precursors. While AKG is primarily known as an intermediate in the citric acid cycle (TCA cycle), it can indirectly contribute to gluconeogenesis through its involvement in amino acid metabolism and energy production. AKG is a key metabolite in the breakdown of glutamine and other amino acids, which can serve as substrates for gluconeogenesis. Additionally, AKG can be converted to oxaloacetate, another TCA cycle intermediate, via anaplerotic reactions, thereby replenishing the cycle and supporting the production of precursors needed for glucose synthesis. This connection highlights AKG's potential to influence gluconeogenesis by maintaining metabolic balance and providing essential intermediates for glucose formation.
| Characteristics | Values |
|---|---|
| Metabolic Pathway | Gluconeogenesis |
| Precursor Molecule | Alpha-Ketoglutarate (α-KG) |
| Conversion Mechanism | α-KG is converted to glucose via a series of enzymatic reactions. |
| Key Enzymes Involved | 1. Glutamate dehydrogenase (reversible) 2. Alanine transaminase 3. Pyruvate carboxylase 4. Phosphoenolpyruvate carboxykinase (PEPCK) 5. Glucose-6-phosphatase |
| Intermediate Steps | 1. α-KG → Glutamate (via glutamate dehydrogenase) 2. Glutamate → Alanine (via alanine transaminase) 3. Alanine → Pyruvate 4. Pyruvate → Oxaloacetate (via pyruvate carboxylase) 5. Oxaloacetate → Phosphoenolpyruvate (PEP) 6. PEP → Glucose-6-phosphate (via PEPCK) 7. Glucose-6-phosphate → Glucose (via glucose-6-phosphatase) |
| Tissue Specificity | Primarily occurs in liver and, to a lesser extent, in kidneys. |
| Energy Requirement | ATP-dependent process, requiring 6 ATP equivalents per glucose molecule. |
| Regulation | Regulated by hormones like glucagon, cortisol, and insulin. |
| Physiological Role | Provides glucose during fasting, low carbohydrate intake, or intense exercise. |
| Clinical Relevance | Important in conditions like diabetes, starvation, and metabolic disorders. |
| Alternative Pathways | α-KG can also enter the citric acid cycle or be used for amino acid synthesis. |
| Reversibility | Most steps are reversible, allowing flexibility in metabolic flux. |
| Substrate Availability | Depends on amino acid availability, particularly glutamate and alanine. |
| pH Dependence | Enzymatic reactions are sensitive to pH, optimal around 7.4. |
| Temperature Dependence | Enzyme activity is temperature-dependent, optimal at 37°C (body temperature). |
| Genetic Influence | Mutations in enzymes like PEPCK or glucose-6-phosphatase can disrupt gluconeogenesis. |
| Nutritional Impact | Dietary protein intake influences α-KG availability for gluconeogenesis. |
Explore related products






What You'll Learn
- AKG to Oxaloacetate Conversion: AKG converts to oxaloacetate via transamination, a key step in gluconeogenesis
- Role of Amino Groups: Amino groups from AKG facilitate the formation of glucose precursors in the pathway
- Mitochondrial to Cytosolic Transport: AKG-derived metabolites move from mitochondria to cytosol for glucose synthesis
- Enzyme Involvement: Glutamate dehydrogenase and transaminases catalyze AKG’s entry into gluconeogenesis
- Energy Requirements: AKG’s conversion requires ATP and reducing equivalents, essential for gluconeogenic processes
![]()
AKG to Oxaloacetate Conversion: AKG converts to oxaloacetate via transamination, a key step in gluconeogenesis
Alpha-ketoglutarate (AKG) plays a significant role in cellular metabolism, particularly in the context of gluconeogenesis, the process by which glucose is synthesized from non-carbohydrate precursors. One of the critical steps in this pathway involves the conversion of AKG to oxaloacetate (OAA), a key intermediate in gluconeogenesis. This conversion is facilitated by the process of transamination, which is essential for the interconversion of amino acids and keto acids. Transamination involves the transfer of an amino group from an amino acid to a keto acid, catalyzed by transaminase enzymes. In the case of AKG, the enzyme glutamate-oxaloacetate transaminase (GOT) plays a pivotal role in this transformation.
The transamination reaction begins with AKG accepting an amino group from glutamate, forming glutamine and oxaloacetate. This reaction is reversible, allowing the cell to regulate the flow of metabolites based on its energy and nutrient needs. The conversion of AKG to OAA is particularly important because OAA is a direct precursor to phosphoenolpyruvate (PEP), another crucial intermediate in gluconeogenesis. PEP is subsequently converted to glucose-6-phosphate and then to glucose, completing the gluconeogenic pathway. Thus, the AKG to OAA conversion via transamination is not only a metabolic link between amino acid and carbohydrate metabolism but also a vital step in maintaining glucose homeostasis.
The enzyme GOT exists in two isoforms: GOT1, primarily found in the cytosol, and GOT2, located in the mitochondria. This compartmentalization allows for the efficient utilization of AKG in both cellular locations, depending on the metabolic demands of the cell. In the mitochondria, AKG is generated as part of the citric acid cycle, while in the cytosol, it can be derived from amino acid catabolism. The ability of GOT to catalyze the transamination of AKG to OAA in both compartments ensures that gluconeogenesis can proceed under various physiological conditions, such as fasting or prolonged exercise, when glucose levels need to be maintained.
Furthermore, the AKG to OAA conversion is tightly regulated to ensure metabolic efficiency. The activity of GOT is influenced by the availability of its substrates (AKG and glutamate) and the concentration of its product (OAA). Additionally, allosteric regulators and hormonal signals, such as insulin and glucagon, modulate the activity of GOT and other enzymes involved in gluconeogenesis. This regulatory network ensures that the conversion of AKG to OAA is synchronized with the overall metabolic state of the cell, preventing wasteful or excessive glucose production.
In summary, the conversion of AKG to oxaloacetate via transamination is a key step in gluconeogenesis, bridging amino acid and carbohydrate metabolism. Catalyzed by glutamate-oxaloacetate transaminase, this reaction generates OAA, a critical intermediate that feeds into the later stages of gluconeogenesis. The compartmentalization of GOT in both the cytosol and mitochondria, along with tight regulatory mechanisms, ensures that this conversion is both efficient and responsive to cellular needs. Understanding this process provides valuable insights into how cells maintain energy balance and adapt to varying nutritional states.
Where to Find Keto Products: In-Store Shopping Guide
You may want to see also
Explore related products






![]()
Role of Amino Groups: Amino groups from AKG facilitate the formation of glucose precursors in the pathway
Alpha-ketoglutarate (AKG) plays a crucial role in gluconeogenesis, the process by which glucose is synthesized from non-carbohydrate precursors. One of the key aspects of this involvement is the role of amino groups derived from AKG in facilitating the formation of glucose precursors. AKG is an intermediate in the citric acid cycle (TCA cycle) and serves as a pivotal molecule in amino acid metabolism. When AKG is transaminated, it donates or accepts amino groups, which are essential for the conversion of certain amino acids into glucose precursors. This transamination process is catalyzed by enzymes such as alanine transaminase and aspartate transaminase, which transfer amino groups to or from AKG, forming glutamate or other amino acids. These amino acids can then be deaminated to provide the carbon skeletons needed for gluconeogenesis.
The amino groups from AKG are particularly important in the context of gluconeogenesis because they enable the interconversion of amino acids that can directly or indirectly contribute to glucose synthesis. For instance, glutamate, formed through transamination with AKG, can be deaminated to form α-ketoglutarate, which re-enters the TCA cycle. Alternatively, glutamate can be converted to α-ketoglutarate and then to oxaloacetate, a direct precursor for glucose synthesis. This pathway ensures that the carbon backbone of AKG is preserved and utilized for gluconeogenesis. The amino groups, once removed, are typically converted to ammonia and eventually to urea for excretion, allowing the carbon skeletons to be repurposed for glucose production.
Another critical step involving amino groups from AKG is the formation of pyruvate, a key intermediate in gluconeogenesis. Through a series of transamination and oxidative deamination reactions, amino acids derived from AKG can be converted into pyruvate. For example, alanine, which is formed by the transamination of pyruvate with glutamate (itself derived from AKG), can be converted back to pyruvate via alanine transaminase. Pyruvate then enters the gluconeogenic pathway, where it is carboxylated to oxaloacetate and eventually converted to glucose. This highlights how the amino groups from AKG indirectly support the generation of glucose precursors by enabling the cycling of carbon skeletons between amino acids and central metabolic intermediates.
Furthermore, the role of amino groups from AKG extends to the regulation of gluconeogenesis through nitrogen balance. Amino acids are both the source and sink of nitrogen in the cell, and AKG acts as a central hub for nitrogen metabolism. By participating in transamination reactions, AKG helps maintain the balance of amino groups, ensuring that excess nitrogen is removed while preserving the carbon skeletons for gluconeogenesis. This regulatory function is vital, as it prevents the accumulation of toxic ammonia and ensures that the cell can efficiently allocate resources to glucose production when needed, such as during fasting or low carbohydrate availability.
In summary, the amino groups from AKG are indispensable in gluconeogenesis as they facilitate the formation and interconversion of glucose precursors. Through transamination reactions, AKG enables the transfer of amino groups, allowing amino acids to be deaminated and their carbon skeletons to be redirected toward glucose synthesis. This process is essential for maintaining metabolic flexibility, particularly under conditions where carbohydrate availability is limited. By acting as a key player in both amino acid metabolism and gluconeogenesis, AKG ensures that the cell can efficiently utilize available resources to meet energy demands.
Sugar-Free Candy on Keto: A Sweet Treat or Diet Pitfall?
You may want to see also
Explore related products





![]()
Mitochondrial to Cytosolic Transport: AKG-derived metabolites move from mitochondria to cytosol for glucose synthesis
Alpha-ketoglutarate (AKG), a key intermediate in the citric acid cycle, plays a significant role in gluconeogenesis, the process by which glucose is synthesized from non-carbohydrate precursors. For AKG-derived metabolites to contribute to gluconeogenesis, they must first be transported from the mitochondria, where the citric acid cycle occurs, to the cytosol, where the majority of gluconeogenic enzymes reside. This mitochondrial-to-cytosolic transport is a critical step that bridges the metabolic pathways of energy production and glucose synthesis. The process involves specific transport mechanisms and metabolic transformations that ensure the availability of gluconeogenic substrates in the cytosol.
The first step in this transport process is the conversion of AKG into metabolites that can exit the mitochondria. AKG is initially converted to oxaloacetate (OAA) via the action of mitochondrial enzymes such as isocitrate dehydrogenase and malate dehydrogenase. OAA is a crucial intermediate that can either be used directly in gluconeogenesis or further metabolized. To facilitate its transport, OAA is reduced to malate by malate dehydrogenase, a reaction that also regenerates NAD+ from NADH, maintaining the redox balance within the mitochondria. Malate, being more permeable to the mitochondrial membrane, is then transported to the cytosol via the malate-aspartate shuttle or the malate-citrate shuttle.
Once in the cytosol, malate is re-oxidized to OAA, providing the substrate for gluconeogenesis. This step is catalyzed by cytosolic malate dehydrogenase, which also reduces NAD+ to NADH, a cofactor that can be used in other cytosolic processes. OAA is then decarboxylated to phosphoenolpyruvate (PEP) by the enzyme phosphoenolpyruvate carboxykinase (PEPCK), a rate-limiting step in gluconeogenesis. PEP is subsequently converted to glucose-6-phosphate (G6P) through a series of enzymatic reactions, ultimately leading to the synthesis of glucose. This cytosolic pathway ensures that the carbon backbone from AKG is effectively utilized for glucose production.
The efficiency of mitochondrial-to-cytosolic transport is tightly regulated to meet the metabolic demands of the cell. For instance, the malate-aspartate shuttle not only transports malate but also facilitates the exchange of aspartate and glutamate, which helps maintain the mitochondrial NAD+/NADH ratio. Similarly, the malate-citrate shuttle involves the transport of citrate, which can be cleaved in the cytosol to regenerate OAA and acetyl-CoA, further supporting gluconeogenesis. These shuttle systems highlight the intricate coordination between mitochondrial and cytosolic metabolism to ensure a steady supply of gluconeogenic precursors.
In summary, the movement of AKG-derived metabolites from the mitochondria to the cytosol is a multifaceted process that involves specific enzymatic conversions and transport mechanisms. The conversion of AKG to malate, its transport to the cytosol, and subsequent re-oxidation to OAA are pivotal steps that link the citric acid cycle to gluconeogenesis. This transport pathway not only ensures the availability of substrates for glucose synthesis but also maintains the metabolic balance between the mitochondria and cytosol. Understanding these mechanisms provides valuable insights into the regulation of gluconeogenesis and its integration with other metabolic pathways.
Can You Enjoy Bell Peppers on a Keto Diet?
You may want to see also
Explore related products






![]()
Enzyme Involvement: Glutamate dehydrogenase and transaminases catalyze AKG’s entry into gluconeogenesis
Alpha-ketoglutarate (AKG), a key intermediate in the citric acid cycle, can enter gluconeogenesis through a series of enzymatic reactions involving glutamate dehydrogenase (GDH) and transaminases. These enzymes play a pivotal role in interconverting AKG with other metabolites, ultimately funneling carbon into the gluconeogenic pathway. The process begins with the conversion of AKG to glutamate, a reaction catalyzed by glutamate dehydrogenase. GDH is a bidirectional enzyme that can either synthesize glutamate from AKG and ammonium or degrade glutamate to AKG, depending on the cellular metabolic state. In the context of gluconeogenesis, GDH primarily operates in the reductive direction, utilizing NADH or NADPH to reduce AKG to glutamate. This step is crucial as it links AKG, a TCA cycle intermediate, to amino acid metabolism, providing a substrate for subsequent transamination reactions.
Transaminases, also known as aminotransferases, are the next set of enzymes critical for AKG's entry into gluconeogenesis. These enzymes catalyze the transfer of an amino group from an amino acid to an α-keto acid, effectively interconverting amino acids and their corresponding keto acids. In this pathway, glutamate, produced by GDH, donates its amino group to α-ketoglutarate, forming pyruvate and another keto acid, such as oxaloacetate, depending on the specific transaminase involved. The transamination of AKG to pyruvate is a direct link to gluconeogenesis, as pyruvate is a key substrate for glucose synthesis. This reaction is typically catalyzed by alanine transaminase (ALT), which transfers the amino group from alanine to AKG, generating pyruvate and glutamate. The glutamate can then re-enter the cycle, being reconverted to AKG by GDH, thus maintaining a steady supply of substrates for gluconeogenesis.
The involvement of transaminases ensures a dynamic equilibrium between amino acids and keto acids, allowing the cell to respond to metabolic demands. For instance, during periods of glucose need, the transamination of AKG to pyruvate is upregulated, providing more substrate for the gluconeogenic pathway. This process is particularly important in the liver and kidneys, where gluconeogenesis is most active. The interplay between GDH and transaminases not only facilitates the entry of AKG into gluconeogenesis but also highlights the integration of amino acid and carbohydrate metabolism. This metabolic flexibility is essential for maintaining energy homeostasis, especially during fasting or low-carbohydrate conditions.
Furthermore, the regulation of these enzymes is tightly controlled to ensure that AKG is directed towards gluconeogenesis when needed. GDH activity, for example, is influenced by the availability of its cofactors (NADH/NAD+ and NADP+/NADPH) and allosteric regulators, such as ADP and GTP, which activate the enzyme, promoting the reductive amination of AKG. Transaminases, on the other hand, are regulated by the concentrations of their substrates and products, ensuring that the transamination reactions proceed in the direction that supports gluconeogenesis. This regulatory network allows the cell to efficiently manage its metabolic resources, directing AKG towards glucose production when energy demands are high.
In summary, the entry of alpha-ketoglutarate into gluconeogenesis is a finely orchestrated process involving glutamate dehydrogenase and transaminases. GDH initiates the pathway by converting AKG to glutamate, which then serves as an amino group donor in transamination reactions. Transaminases, particularly ALT, catalyze the transfer of the amino group from glutamate to AKG, producing pyruvate, a direct precursor for glucose synthesis. This enzymatic cascade not only provides a metabolic link between the TCA cycle and gluconeogenesis but also exemplifies the intricate regulation of cellular metabolism to meet energy requirements. Understanding these enzyme-driven processes is crucial for comprehending how cells adapt to varying nutritional states and maintain metabolic balance.
Are Atkins Bars Keto-Friendly? A Guide to Low-Carb Snacking
You may want to see also
Explore related products






![]()
Energy Requirements: AKG’s conversion requires ATP and reducing equivalents, essential for gluconeogenic processes
Alpha-ketoglutarate (AKG) plays a crucial role in cellular metabolism, particularly in the context of gluconeogenesis, the process by which glucose is synthesized from non-carbohydrate precursors. For AKG to contribute to gluconeogenesis, it must undergo a series of metabolic conversions that require significant energy input. Central to these conversions are the requirements for ATP (adenosine triphosphate) and reducing equivalents, such as NADH (nicotinamide adenine dinucleotide) and NADPH (reduced nicotinamide adenine dinucleotide phosphate). These molecules are essential for driving the enzymatic reactions that transform AKG into intermediates that can enter the gluconeogenic pathway.
The first step in AKG's conversion involves its transformation into oxaloacetate, a key intermediate in the gluconeogenic process. This reaction is catalyzed by the enzyme mitochondrial oxoglutarate carboxylase, which requires ATP as an energy source. The consumption of ATP highlights the energy-intensive nature of this step, underscoring the importance of cellular energy reserves in supporting gluconeogenesis. Without sufficient ATP, the conversion of AKG to oxaloacetate would be impaired, limiting the availability of substrates for glucose synthesis.
Following the formation of oxaloacetate, further metabolic steps are necessary to integrate this intermediate into the gluconeogenic pathway. These steps often involve the reduction of carbonyl groups, a process that relies on reducing equivalents like NADH and NADPH. For instance, the conversion of oxaloacetate to malate, a precursor in gluconeogenesis, requires NADH as a cofactor. Similarly, subsequent reactions in the pathway, such as the reduction of pyruvate to lactate or the regeneration of oxaloacetate from phosphoenolpyruvate, also depend on these reducing equivalents. The availability of NADH and NADPH is therefore critical, as they provide the electrons needed to drive these reductive reactions forward.
The interplay between ATP and reducing equivalents in AKG's conversion to gluconeogenic intermediates reflects the broader metabolic coordination required for energy homeostasis. ATP serves as the primary energy currency, powering the endergonic reactions that would otherwise be non-spontaneous. Meanwhile, reducing equivalents act as electron carriers, facilitating the transfer of electrons in redox reactions that are integral to metabolic transformations. Together, these molecules ensure that the energy demands of gluconeogenesis are met, even under conditions of carbohydrate scarcity.
In summary, the conversion of AKG into gluconeogenic intermediates is a highly energy-dependent process that relies on both ATP and reducing equivalents. These requirements underscore the metabolic cost of synthesizing glucose from non-carbohydrate sources and highlight the importance of cellular energy management in maintaining metabolic flexibility. Understanding these energy requirements provides insights into how cells prioritize and allocate resources to support essential physiological processes, such as gluconeogenesis, in response to varying nutritional and metabolic states.
Freezing Cooked Keto Mozzarella Sticks: Tips for Perfect Storage
You may want to see also
Frequently asked questions
Alpha-ketoglutarate (AKG) is an intermediate in the citric acid cycle (Krebs cycle). While it is not a direct precursor for gluconeogenesis, it can contribute indirectly by replenishing oxaloacetate (OAA) through anaplerotic reactions, which supports the gluconeogenic pathway.
No, alpha-ketoglutarate cannot be directly converted into glucose. However, it can be converted into oxaloacetate via transamination or other metabolic pathways, which can then enter gluconeogenesis.
Alpha-ketoglutarate can be transaminated to form aspartate, which is then converted to oxaloacetate. This oxaloacetate can then be used in the gluconeogenic pathway to produce glucose.
The conversion involves transaminases (e.g., glutamate-oxaloacetate transaminase) and other enzymes like aspartate aminotransferase, which facilitate the transformation of alpha-ketoglutarate to oxaloacetate via intermediate steps.
Alpha-ketoglutarate plays a secondary role in gluconeogenesis. It is not a primary substrate like pyruvate, lactate, or glycerol, but it supports the pathway by maintaining the levels of key intermediates like oxaloacetate.































